トップページ > 研究組織一覧 > 分野・独立ユニットグループ > がんRNA研究分野 > 研究プロジェクト > がんにおけるRNAスプライシング異常
がんにおけるRNAスプライシング異常
更新日 : 2024年3月22日
SF3B1変異による発がん機構の解明
SF3B1は、がんの中で最も高頻度に遺伝子変異が見つかるスプライシング因子です。
我々は変異型SF3B1による発がん機序について、全がんスプライシング解析やマウスモデル、分子生物学的手法、臨床検体を用いた検証などを駆使して取り組んできました。
その結果、変異型SF3B1はアレル特異的、かつ組織特異的にスプライシング異常をきたすこと、慢性リンパ性白血病においてはMYCシグナルを更新させること(図)、MYC活性化の原因が変異型SF3B1によるPPP2R5Aのミススプライシングによるものであること、PPP2R5A異常はBCL2も同時に活性化すること、PP2A賦活剤がSF3B1変異白血病モデルに選択的に有効であることを最近報告しました(Liu Z*, Yoshimi A*,#, et al. Cancer Discov. 2020)。
PPP2R5Aはプロテインフォスファターゼ2(PP2A)のサブユニットの一つであり、そのミススプライシングはSF3B1変異白血病において非常に強く生じていますが、PPP2R5Aの機能についてはまだ十分に解明されていません。
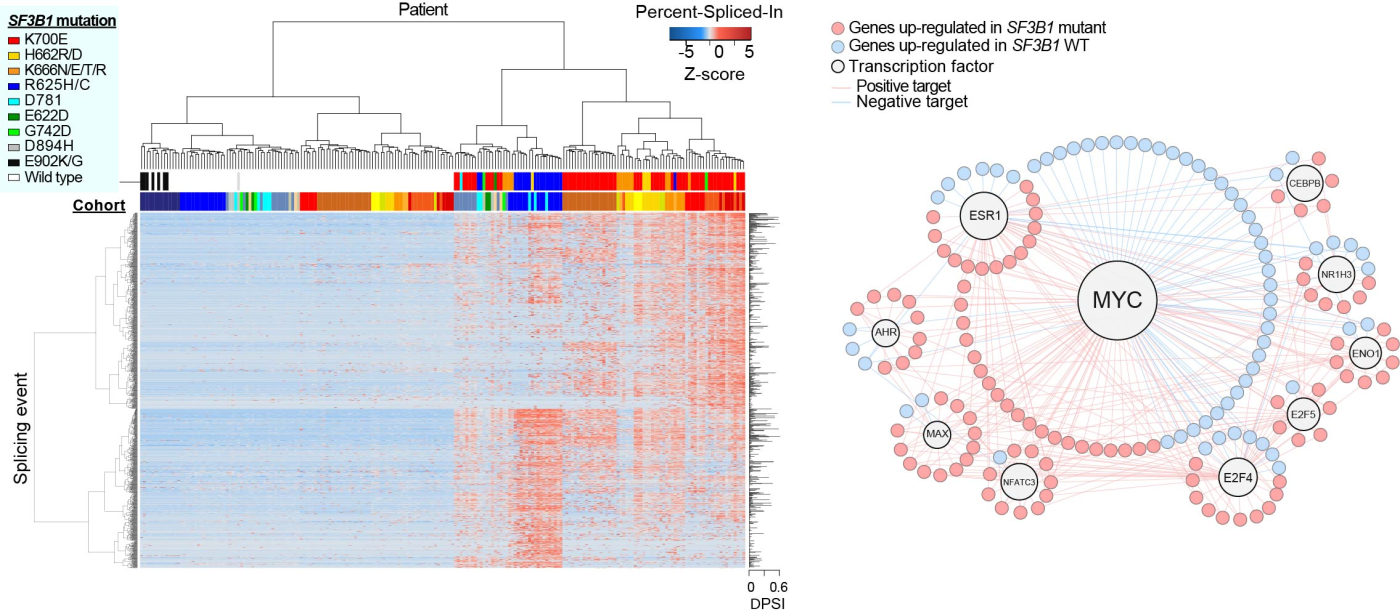
図:全がんスプライシング解析により、SF3B1変異はホットスポットやがん種によって異なるスプライシング異常を来すことが分かった(左)。慢性リンパ性白血病においてはSF3B1変異がMYCの活性化を誘導する(右)。
PPP2R5AはPP2A複合体の中でもユニークな特徴があり、我々の研究室では、PPP2R5Aの発がんにおける機能を解明し、新たな治療標的を同定することを目指しています。また、PPP2R5AのBRET(bioluminescent resonance energy transfer)センサーの開発にも取り組んでいます。
主担当者:山内 浩文
Grant: ASH Global Research Award
HireCLIP-seqを用いたスプライシング制御機構の解明
そもそも、スプライシングはどのように制御されているのでしょうか?
もちろん、スプライシングという現象が1997年に発見されて以来、様々な研究が進められてきました。
一方で、未だにメジャーなスプライシング因子(SF)であるSRタンパク質ファミリーやhnRNPタンパク質ファミリーが、RNA上のどこに結合してRNAをどう制御しているのかは、明らかになっていません(例えば、356種類ものRNA結合蛋白質の網羅的プロファイリングを謳うENCODE projectにおいても、結合部位が正確に(D-score < 0.05)定義されたSFは16種類しかありません)。
これは、SFのRNA結合部位の解析方法がすべて抗体依存的であることに起因します(つまり、親和性・特異性に優れた抗体が買えなければ解析不能です)。
そこで、わたしたちは、注目するSFに対する抗体が入手可能か否かにかかわらず、高精度にRNA結合部位を同定可能な(D-score<1.0e-4)、Highly reproducible single-end enhanced cross-linking and immunoprecipitation sequence (HireCLIP-seq) 法を開発しました。
今まで得ることできなかった貴重なデータに基づき、RNAスプライシングやRNA修飾とのOmics解析により、今までに見えなかったスプライシング制御機構を知ることが可能になりました。
主担当者:吉田 澪奈
- がんRNA研究分野
- 研究プロジェクト
- RNAとは?【がんRNA研究分野】
- 業績論文
- 著書
- 研究室メンバー【がんRNA研究分野】
- コンタクト

